〜分子機構と生理的意義〜
(新潟大学大学院医歯学総合研究科)
共同研究者:栗原 悠介
(九州大学大学院医学系学府)
■研究の背景
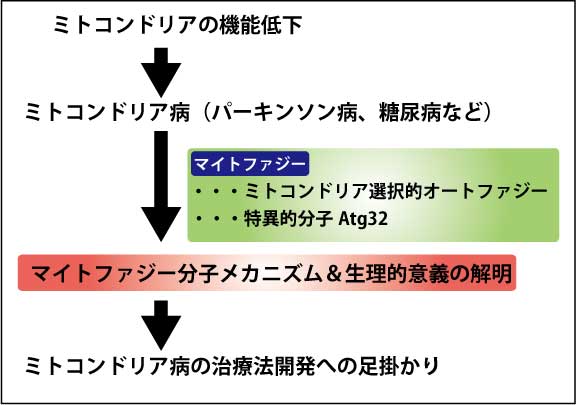
ミトコンドリアは細胞が必要とするエネルギーの大半を供給する重要なオルガネラであると同時に、活性酸素を大量に放出する場でもある。そのため、ミトコンドリアの品質と量を維持することは、細胞が恒常性を維持するために必要不可欠である。マイトファジーとはミトンドリアを選択的に分解する現象であり、ミトコンドリアの品質と量の管理に重要だと考えられている。近年、様々な神経疾患が異常なミトコンドリアの蓄積によるものだと考えられていることから、マイトファジーの理解を深める事は、神経疾患の治療系の開発に繋がると期待している。
■研究の目的
マイトファジーの理解を深めるため、出芽酵母を用いて、マイトファジーの分子メカニズム及び生理的意義の解明を目的とした。
■分子メカニズム
拡大図はこちら
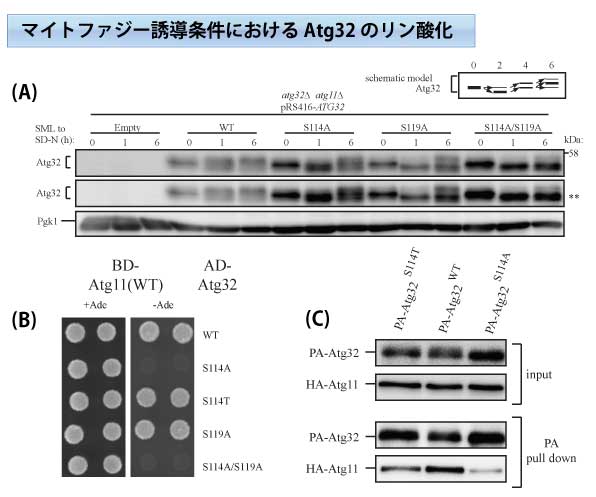
下図に示すように、Atg32はミトコンドリア外膜に局在するマイトファジーの特異的因子である。(A)マイトファジー誘導条件(栄養飢餓:SD-N)に伴い、Atg32のアミノ基末端から114番目と119番目のセリン残基のリン酸化が確認できた。(B), (C) イーストツーハイブリット法、プルダウンアッセイ法により、114番目のリン酸化されたセリン残基を介して、アダプター蛋白質であるAtg11がAtg32と結合することが明らかとなった。
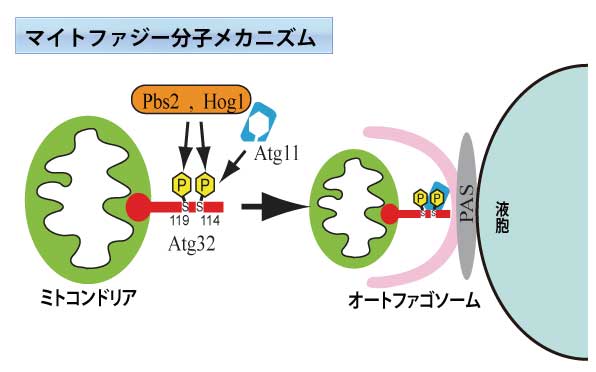
細胞が栄養飢餓状況に置かれると、浸透圧感受性のキナーゼであるPbs2やHog1を介して、Atg32はリン酸化される。次にリン酸化された114番目のセリン残基を介して、Atg32はAtg11と結合し、ミトコンドリアはオートファゴソーム形成の場であるPAS(Pre-Autophagosome-Site)領域へと運ばれる。
■生理的意義
拡大図はこちら
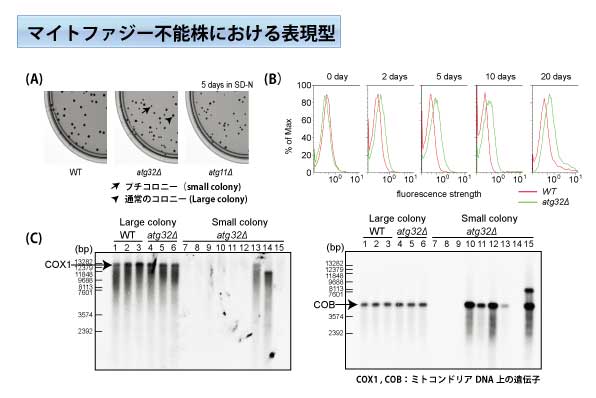
(A)マイトファジー不能株(atg32Δ,atg11Δ)では、生育不良の結果生じるプチコロニーが確認できる。(B)フローサイトメトリーの解析結果より、栄養飢餓条件において、マイトファジー不能株からの活性酸素産出量は野生株と比較して増加する事が確認できた。(C)サザンブロット解析の結果から、マイトファジー不能株のミトコンドリアDNAは、栄養飢餓条件において欠損(あるいは一部欠失)することが明らかとなった。
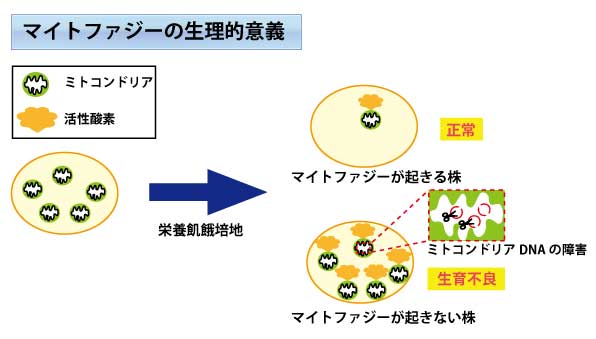
栄養飢餓条件において、野生株は余分なミトコンドリアを排除することが可能であるが、マイトファジー不能株はミトコンドリアを排除することができず必要以上の活性酸素が産出されてしまう。多量の活性酸素はミトコンドリアDNAに障害を与え、その結果、生育不良になってしまう。
研究課題:ミトコンドリアオートファジーの基礎研究
研究組織:医・医病 審査部門:理工農系 採択年度:H23 整理番号:23322
種 目:D-1タイプ( 萌芽的若手研究)
代表者:神吉 智丈(九州大学病院 助教 現:新潟大学 教授)
フレームが表示されていない場合はこちら